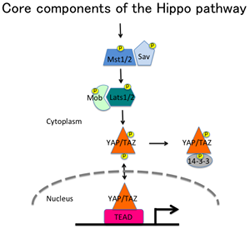
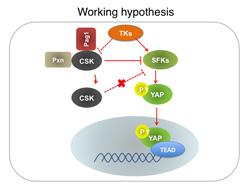
がんは、細胞の増殖や生存を制御するシグナル伝達機構の破綻によって生じることが知られています。しかし、一部のがんを除いて発がんのメカニズムの詳細は解明されていません。近年、多くのがんでHippo経路というシグナル伝達経路に異常があることが明らかにされました。われわれは、1990年代に白血病と関わりのある新規のタンパク質リン酸化酵素Kpm (LATS2)を同定していましたが、それが、Hippo経路の中核分子の1つであることが判明しました(右上図)。Hippo経路は、最終的なエフェクター分子であるYAP/TAZの活性化を制御するセリン/スレオニンのリン酸化カスケードと見なすことができます。YAP/TAZの活性化は多くのがんで見られます。最近では、Hippo経路の制御とは別に、Srcファミリーリン酸化酵素 (SFKs) によるYAPのチロシンリン酸化もYAPの活性化を引き起こすことが分かっています(右下図)。われわれは、これらのことを考慮しながら、主にYAPの活性化機構について解析しています。
DNA損傷による有害な現象を未然に防ぐ為に、生物はDNA修復機構によりDNA損傷をゲノムから除去しています。DNA損傷のなかでも最も重篤な損傷の一つであるDNA二重鎖切断(DNA double-strand breaks: DSBs)は主に非相同末端再結合あるいは相同組換えにより修復されます。西助教を中心に、これらの修復経路において重要な役割を果たすタンパク質翻訳後修飾の一種であるユビキチン化に着目し、DSB修復の詳細な制御機構を分子レベルで明らかにすることを目標として研究を進めています。