~ 形が崩れても機能は残る ~ 網膜の層構造形成を制御する因子と、神経系の構造-機能間の新たな関係を発見
立命館大学総合科学技術研究機構・システム視覚科学研究センターの上野明希子助教(薬学部)、立花政夫上席研究員、作田木南(薬学研究科2回生)、小池千恵子教授(薬学部・センター長)らの研究チームは、細胞接着因子Afadinが神経網膜の層構造およびシナプス形成、細胞分化に必須であることを明らかにしました。さらに、Afadinを欠損し網膜層構造が大きく崩壊したマウスでも、視覚機能の一部が保持されることを明らかにしました。
本研究成果は、2025年12月23日付で、国際科学雑誌「eLife」に掲載されました。
本件のポイント
- 細胞接着因子Afadinが網膜の層構造および視細胞シナプス形成に必須であることを発見
- 網膜の層構造形成に発生段階のアドへレンスジャンクション(AJ)が寄与していることを発見
- 網膜の層構造および視細胞シナプスが大きく崩壊しても、一部の視覚情報処理が維持されることを確認
研究成果の概要
視覚形成の第一段階を担う神経網膜は整然とした層構造を形成しています。網膜の層構造は視覚形成において重要な役割を担うと考えられてきましたが、層構造の破綻が網膜および視覚機能に与える影響や、層構造形成の制御機構はほとんど解明されていませんでした。
本研究では、細胞接着因子で、アドへレンスジャンクション(AJ)※1の主要構成因子であるAfadinに着目し、網膜特異的にAfadinを欠損させたマウス(Afadin cKOマウス)※2を用いて組織学的解析、電気生理学的解析、および視覚行動解析を行いました。Afadin cKOマウス網膜では顕著な層構造崩壊、視細胞シナプス※3の大幅な減少、網膜細胞の運命決定異常が認められ、発生段階で異所的なAJと網膜先駆細胞※4の位置異常が確認されました。一方で、電気生理学的解析および視覚行動解析の結果、Afadin cKOマウスは網膜機能を部分的に保持しており、物体の動きを認識できることが示されました。これらの結果は細胞接着因子Afadinが網膜の層構造形成、シナプス形成、および神経細胞分化に必須であり、発生段階のAJが網膜前駆細胞の移動と分化を正常に制御することで層構造形成に寄与する可能性を示唆しています。また、網膜層構造やシナプスに重大な異常が生じた場合でも、網膜神経回路の機能が一部維持され、視覚機能も残り得ることを示しています。
本研究は、網膜の形態形成に関する理解を深めるとともに、神経組織の「形態と機能の関係」に関する新たな知見を提供します。
研究の背景
私たちの視覚形成には目の奥にある網膜が欠かせない役割を担っています。目に入った光は網膜の視細胞で電気信号に変換され、その後、複雑な情報処理を経て網膜神経節細胞(RGC)から脳へとシグナルが伝達されます。これにより、私たちは“物が見えた”と認識します。
網膜は視細胞・双極細胞・神経節細胞などの6種類の神経細胞と1種類のグリア細胞から主に構成されています(図1)。これらの神経細胞が適切な場所で適切な相手とシナプスを介して接続することで、精密な神経回路と層構造を形成されます。
この層構造は大きく外顆粒層(ONL)、外網状層(OPL)、内顆粒層(INL)、内網状層(IPL)、神経節細胞層(GCL)に分けられ、各層ごとに構成成分が異なります(図1)。網膜の層構造は視覚機能において重要であると考えられており、層構造が種を超えて保存されていることからもその重要性が伺えます。
しかし、この層構造形成の分子機構や、層構造が破綻した場合の網膜および視覚機能への影響は報告されていませんでした。
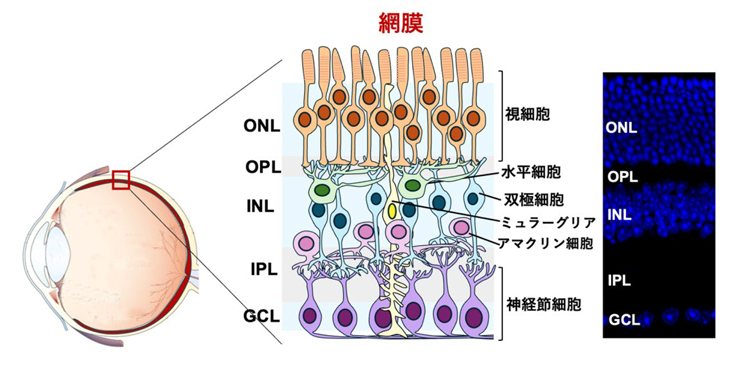 図1:網膜の構造
図1:網膜の構造
(左)網膜の構造図。眼球内の赤い部分は網膜を表す。(右)成体マウス網膜切片のDAPI染色像。
研究内容
本研究ではまず、Afadin cKOマウスを用いて組織学的解析を行いました。その結果、成体のAfadin cKOマウス網膜では外側(視細胞側)の層構造が顕著に崩壊し、特にONLとOPLは判別できない状態でした。一方、内側(神経節細胞側)の層構造は比較的保たれていましたが、各網膜細胞の分布に異常が確認されました (図2)。
また、Afadin cKO網膜では視細胞の外節※5の位置と形態にも異常が見られ、視細胞-双極細胞間のシナプスも著しく減少していました(図2)。また、Afadin cKO網膜では視細胞が有意に減少し、双極細胞が有意に増加していました。
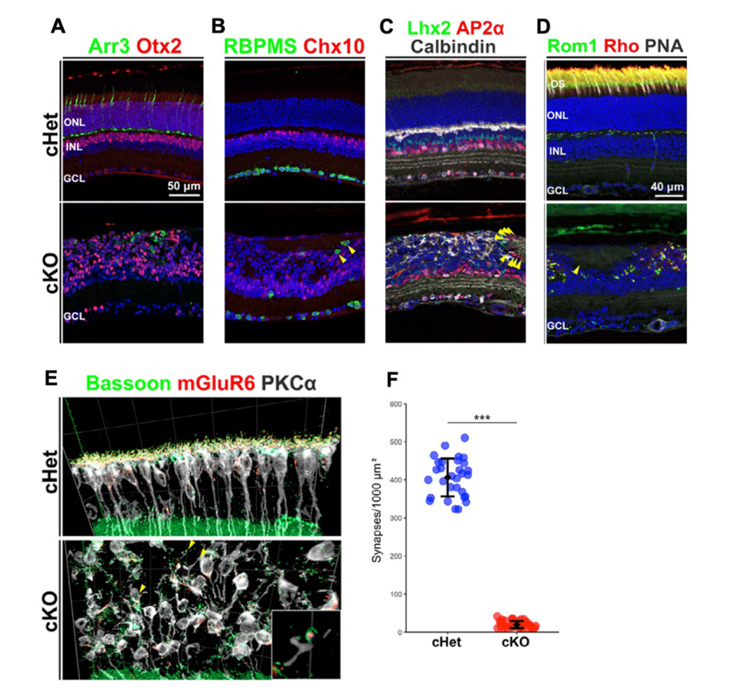 図2:Afadin cKOマウス網膜の組織解析結果
図2:Afadin cKOマウス網膜の組織解析結果
(A-E)コントロールマウス(cHet)とAfadin cKOマウス網膜の免疫組織染色像。Arr3:錐体視細胞マーカー,Otx2:視細胞と双極細胞のマーカー,RBPMS:RGCマーカー,Chx10:双極細胞マーカー,Lhx2:ミュラーグリアマーカー,AP2α:アマクリン細胞マーカー,Calbindin:水平細胞と一部のアマクリン細胞マーカー,Rom1とRho:rodマーカー,PNA:錐体視細胞マーカー,BassoonとmGluR6:シナプスマーカー,PKCα:双極細胞マーカー。(E)は3次元構築図。(F)網膜表面1mm2直下に存在する視細胞シナプスの数。
Afadin cKOマウス網膜の機能を評価するため、多電極アレイ(multi-electrode array, MEA)※6を用いて電気生理学的解析を行いました。その結果、Afadin cKO網膜では顕著な組織学的異常にもかかわらず、光刺激に対するRGCの応答が記録されました。
コントロール網膜のRGCはその応答特性から5種類に分類されましたが、そのうち2種類はAfadin cKO網膜では確認されませんでした。さらに、Afadin cKO網膜で一部のRGCが受容野※7を持つことを示唆するデータが得られました(図3)。これらの結果はAfadin cKOマウスでは網膜の機能が部分的に維持されている可能性を示しています。
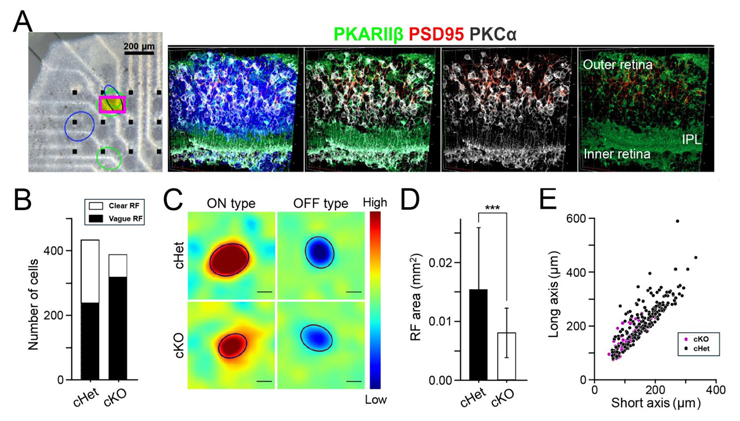 図3:Afadin cKOマウス網膜の電気生理学的解析結果
図3:Afadin cKOマウス網膜の電気生理学的解析結果
(A)MEA上のAfadin cKOマウス網膜と受容野が検出された部位(楕円)(左)。四角で囲った受容野検出部位を解析後組織解析し、構造が異常となっていることを確認した(右4つ)。(B-E)コントロールマウス(cHet)とAfadin cKOマウス網膜の受容野の数(B)、検出された受容野の例(C)、受容野の大きさ(D)、受容野の長軸と短軸の長さ(E)。
Afadin cKOマウス網膜からは受容野が検出され、受容野検出部位であっても著しい構造異常が見られることを示す。
MEA解析の結果を受け、Afadin cKOマウスの視覚機能を個体レベルで評価するために、視運動反応(optomotor response, OMR)試験※8を行いました。Afadin cKOマウスが視覚刺激に対して追従行動をとるようすが確認されましたが、その応答回数は正常なマウスに比べて有意に減少していました(図4)。
また、比較的高い空間周波数の刺激に対しては、ほとんど追従行動を示しませんでした(図4)。
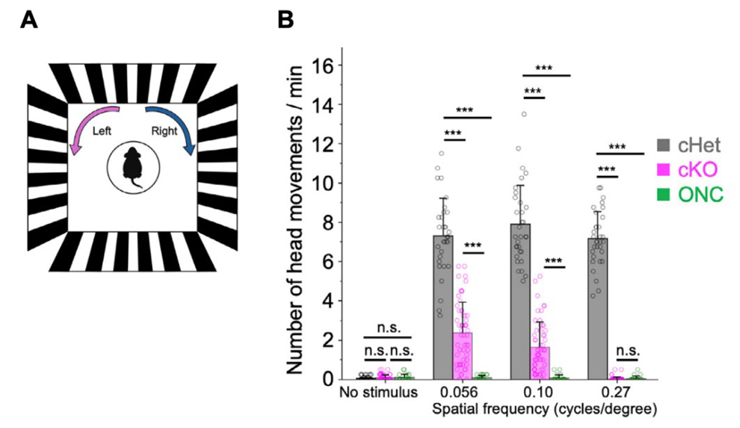 図4:Afadin cKOマウスを用いたOMR試験の結果
図4:Afadin cKOマウスを用いたOMR試験の結果
(A)OMR試験の概略図。マウスに右または左に動く縞模様の刺激を提示し、追従行動回数を測定する。(B)コントロールマウス(cHet)、Afadin cKOマウス、視神経挫滅マウスのOMR試験結果。横軸の0.056,0.1,0.27は空間周波数(縞模様の太さ、高いほど細い)を表す。Afadin cKOマウスは空間周波数0.056,0.1の刺激に対して追従運動を示すが、コントロールマウスに比べて頻度が少なく、空間周波数0.27の刺激では追従運動を示さない。視神経挫滅マウスは全ての視覚刺激に応答を示さず、全てのマウスは刺激なしでは追従運動を示さない。
これらの追従運動は視神経挫滅により外科的に視覚を剥奪したマウスでは観察されませんでした。これらの結果は、Afadin cKOマウスは正常なマウスに比べて精度は低いものの、視覚刺激を認識していることを示唆しており、一部視覚機能が維持されている可能性が考えられます。
研究グループはさらに、発生段階のAfadin cKOマウス網膜において、異所的なAJが形成されることを発見しました。また、Afadin cKO網膜では網膜前駆細胞の分布パターンが変化しており、正常な網膜に比べて分布範囲が有意に拡大していることも確認されました。
本研究結果から、Afadinは網膜の層構造形成、シナプス形成、正常な細胞運命決定に必須であり、Afadin cKO網膜では異所的なAJを足場とした細胞移動の結果、網膜層構造やシナプス形成に異常が生じる可能性が示唆されました。また、Afadin cKOマウス網膜では顕著な層構造の崩壊、視細胞シナプスの減少、外節の異常が生じているにもかかわらず、このマウスの網膜および視覚機能が部分的に維持されている可能性が示されました。
社会的な意義
私たちが外界から得る情報の約8割は視覚に由来するといわれており、視覚機能の障害はときに著しい生活の質(QOL)の低下を招きます。視覚形成メカニズムの理解と解明は、視覚障害の治療法の開発や改良において重要ですが、完全な解明には至っていません。特に、組織学的に異常がある網膜がどの程度機能を保持し得るのかという点について、ほとんど報告がありませんでした。
本研究結果は、この未解明だった点に関して新たな知見を提供するものです。また、本研究は脳を含む中枢神経系の層構造と機能の関係を解明するうえで、1つの重要な知見になり得ると考えられます。
さらに、再生医療の臨床試験において、失明疾患患者の網膜にiPS細胞由来の視細胞シートを移植する手術が行われていますが、移植後一時的に視力を回復した例があります。しかしながら、視細胞シート移植後の網膜では視細胞層は整列しません。本研究は、網膜層構造が正常化しなくとも、一部機能が回復すれば視力を発揮できることに対する説明となると考えています。
研究者のコメント
構造が崩壊した網膜でも機能が一部維持されているのは想定外でした。なぜ機能が維持されるかについては不明な点が多く、今後の課題かと思います。本研究が今後の神経系の形と機能の関係性の研究発展に少しでも貢献できれば幸いです。これまでサポートいただいた全ての方と、研究遂行の助成予算を支出いただいた方々に心より感謝申し上げます。
共同研究先
本研究は、三好淳先生(元大阪成人病センター)、高井義美教授(神戸大学)との共同研究です。
論文情報
- 論文名:Afadin-deficient mouse retinas exhibit severe neuronal lamination defects but preserve visual functions
- 著者:Akiko Ueno, Konan Sakuta, Hiroki Ono, Aki Hashio, Haruki Tokumoto, Mikiya Watanabe, Taketo Nishimoto, Toru Konishi, Yuki Emori, Shunsuke Mizuno, Mao Hiratsuka, Jun Miyoshi, Yoshimi Takai, Masao Tachibana, Chieko Koike
- 発表雑誌:eLife
- 掲載日:2025年12月23日 0時0分(日本時間)
- DOI:https://doi.org/10.7554/eLife.105627.3
- URL:https://elifesciences.org/articles/105627#content
用語説明
- ※1 アドへレンスジャンクション(AJ)細胞間接合部位で形成される、タンパク質複合体からなる機能構造体。主に物理的に強固な細胞間接着を担い、発生段階の細胞移動や生体の恒常性に大きく寄与します。
- ※2 Afadin cKOマウスCre-loxPシステムによって網膜特異的にAfadinを欠損したマウス。Afadinflox/floxマウスとDkk3-Creマウスを交配により作製しました。
- ※3 シナプス神経細胞間や神経細胞と他の細胞の間で形成される接着構造で、情報の伝達が行われる場所。信号を出力する細胞と信号を受け取る細胞の接触部位で形成され、神経系における情報伝達の基盤となります。
- ※4 網膜先駆細胞細胞分裂能を有し、限定された種類の網膜細胞に分化できる細胞。
- ※5 外節視細胞の小器官の1つで、光を電気信号に変換する場所。繊毛が発達した構造で、外節内には大量の光受容タンパク質が埋め込まれた膜が積み重なっています。視細胞が光を受け取って電気信号に変換するうえで重要な役割を担います。
- ※6 多電極アレイ(multi-electrode array,MEA)複数の微小な電極を規則的に配置した装置で、神経組織などの電気的な活動を同時に多点で記録できる技術です。
- ※7 受容野感覚処理系の個々の神経細胞(本研究ではRGC)が、外界からの刺激に対して反応することのできる空間範囲。
- ※8 視運動反応(optomotor response,OMR)試験横に動く縞模様を動物の前に提示し、縞模様の動きに対する追従行動回数を計測する解析方法。縞模様の太さ(空間周波数)や動く速さ(時間周波数)を変更して解析することが多い。正常なマウスは一定の空間および時間周波数で追従運動を示す。マウスやラットの視覚機能の評価に使用される。
関連情報



<世界から立命館へ!> 研究室だけじゃない—奈良での生活、AI、ロボティクスについて:エル・ハフィ・ロトフィ先生
